Dr. Angela Rasmussen: Stop Using Libel, Lies, and Rhetoric to Harass and Defame Me
I request that you cease and desist

I am currently trying to deal with a very serious health crisis in my family. Despite this, there are academics who remain doggedly determined to smear my professional reputation by publicly identifying me alongside accusations that are outright lies. The success of an academic’s career correlates with their professional reputation. Consequently, I have the right to defend my reputation when others choose to publicly attack me.
Enter Dr. Angela Rasmussen, who is apparently a virologist at the University of Saskatchewan. I had no idea who she was until members of the public recently made me aware that she made a public post about me in lieu of talking to me directly about concerns that she obviously has. I am sick and tired of being mistreated by these so-called professionals that fail to demonstrate basic courtesies like talking to those that they take issue with before publicly airing their ill-conceived assumptions. Since Dr. Rasmussen decided to engage by making public declarations about me, I will respond in public fashion via this open letter to compensate for the harms she has and is causing. I will email a link to this article to her.
This is what Dr. Rasmussen posted about me…
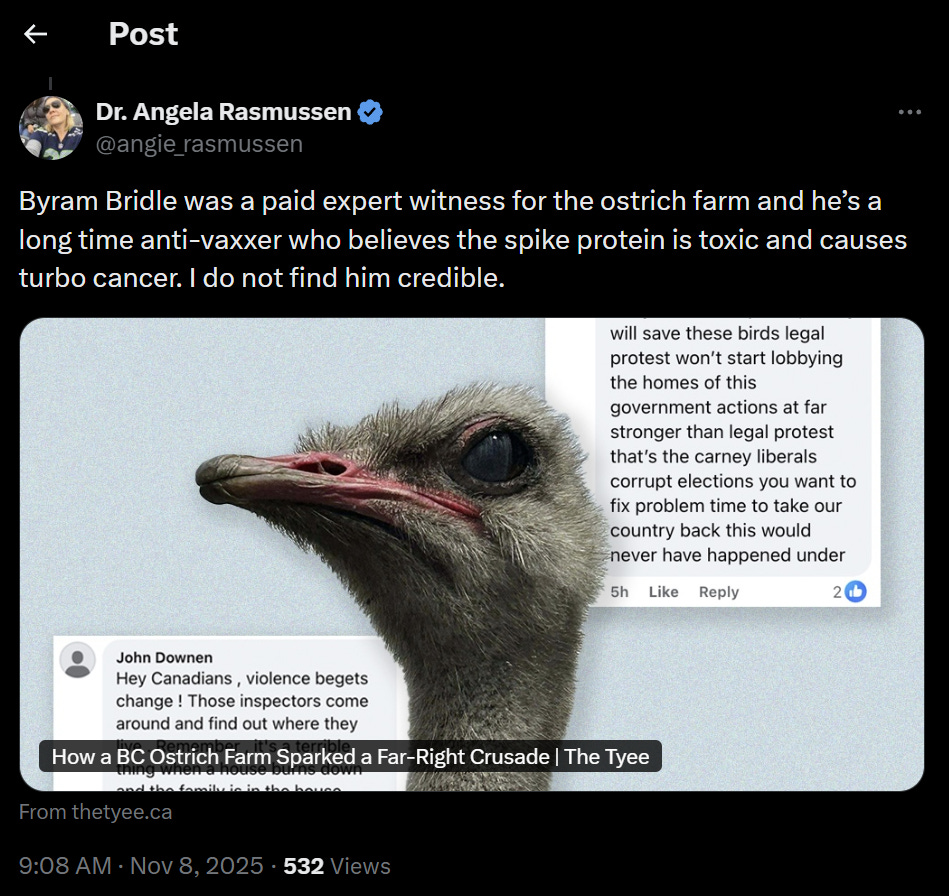
Evidence of a disrespectful nature
“Dr. Angela Rasmussen“ versus “Byram Bridle”
Dr. Rasmussen, I noted that you refer to yourself as “Dr. Angela Rasmussen” but to me to me as “Byram Bridle”, having dropped my professional title of “Dr.”. This reflects extremely poorly on you as it shows disrespect and immaturity. Disrespect is not a way to promote constructive discussions of science. This was not a good start.
A blatant lie
“Byram Bridle was a paid expert witness”
Dr. Rasmussen, you claimed that I served as a paid expert witness. This is a strategy that is often used to try to mislead the public into thinking that an expert is somehow inherently biased in favour of those who pay them. What you failed to disclose is that professionals are entitled and often expected to receive payment for their professional services in court. Also, experts are sworn to objectively serve the interests of the court, not any of the litigants. Indeed, Dr. Rasmussen, you failed to disclose that I placed myself under oath and swore to provide objective expert testimony in service of the court, not the owners of the ostrich farm. I was expected to back up any scientific claims with evidence, which I did in a very robust fashion, ensuring that the court was accurately educated on all relevant points based on the overall weight of the scientific evidence. Most notably, I was not paid! Claiming that an unpaid witness was paid is, quite simply, a lie and liars cannot be trusted. Anything that a liar said prior and anything they say after cannot be trusted. The onus then falls on those who lie publicly to attempt to earn trust back from the public. And there can be no expectations placed on the public to reinstate their trust in the liar.
Failure to vet sources of information
Dr. Rasmussen, your single reference to try to support your claim that I was a paid expert in the ostrich trial is a clearly biased hit-piece published in The Tyee, which makes no claim that I was a paid witness. An academic should actually read their references and know what they do and do not support. This does not speak well of an academic’s competence.
Defamation through libel
“He’s a long-time anti-vaxxer”
I am literally a vaccinologist. I specialize in vaccinology and have served the public in this field for most of my scientific career. I have published and taught extensively in this area and contend that I likely have superior expertise in this area than what you have, Dr. Rasmussen, although this is based on my limited perusal of your qualifications. Put another way, Dr. Rasmussen, I would be happy to debate you anytime on the topic of vaccines, ideally in a public forum, to allow a real-time assessment of who holds superior qualifications in this sub-discipline of immunology. So, this is literal defamation expressed through libel. I don’t know how an academic professional gets away with this.
Rhetorical rubbish
Applying labels with negative implications to people that you don’t know and in an absence of evidence is the hallmark of harmful rhetoric. Any claims made by people who purport to be combatants of misinformation, only to resort to rhetoric are to be discounted. It is usually a sign of someone who lacks professionalism and/or expertise in the subject matter.
Follow the science, not opinions lacking evidence
“Who believes the spike protein is toxic”
This is interesting. Dr. Rasmussen, you quickly diverged from the current topic of the ostrich case to what is obviously a historical grievance of yours that you apparently cannot let go of. Clearly, you must have disliked statements that I made historically about the spike protein, despite them being demonstrably true based on the overwhelming peer-reviewed scientific evidence. All I have ever done in my public messaging about the spike protein is interpret objective scientific facts so that laypeople can understand them. I do this to facilitate informed consent. I have never relayed beliefs about the spike protein, only objective truths.
Dr. Rasmussen, you seem to be implying that you do not recognize any potential harmful bioactivity that the spike protein may mediate inside the human body. This suggests that you are unaware of the plethora of literature that clearly demonstrated the central role that the spike protein or its subunits played in the context of natural infections when they got distributed systemically through viremia (meaning when the virus got spread throughout the body via the bloodstream). There are lots of publications on this topic of the natural pathophysiology of the virus.
Dr. Rasmussen, your statement also implies that you are unaware of the fact that spike encoded by the vaccines is fully capable of binding to the same receptors as the native protein, and therefore, retaining the potential to mediate the same harms as the native protein. In fact, the spike proteins used for the vaccines were specifically selected for their ability to strongly bind to the ACE-2 receptor because this was hypothesized to be necessary to generate the best neutralizing antibodies (i.e., ones that could bind directly to and, therefore, block the receptor-binding domain). This strategy was published by the scientists that designed the vaccines.
Dr. Rasmussen, your statement further implies a lack of understanding of the plethora of data demonstrating the systemic biodistribution of the lipid nanoparticles that carry the modified RNAs encoding the spike protein, which aligns with the massive body of literature showing this throughout the history of development of this technology.
And finally, Dr. Rasmussen, your statement implies that you are unaware of the literature directly showing a wide variety of harmful bioactivities mediated specifically by vaccine-encoded spike proteins.
So, Dr. Rasmussen, to correct your ignorance on these matters, and in stark contrast to your evidence-free statement, here is some literature that I recommend you peruse to become better informed on these subjects:
Studies demonstrating the toxic nature of the spike protein, whether it be following natural infections or vaccination:
SARS-CoV-2 Spike Protein Impairs Endothelial Function via Downregulation of ACE2, Lei Y, et al., Circulation Research, 2021, doi:10.1161/CIRCRESAHA.121.318902. Spike protein alone damages vascular endothelial cells by downregulating ACE2, impairing mitochondrial function, reducing eNOS activity, and increasing glycolysis, leading to endotheliitis observed in severe COVID-19.
Sustained Vascular Inflammatory Effects of SARS-CoV-2 Spike Protein on Human Endothelial Cells, Cuevas S, et al., Inflammation, 2025, PMID: 39739157. Spike protein triggers prolonged ICAM1 expression, leukocyte adhesion, and procoagulant state in human aortic and pulmonary microvascular endothelial cells under flow conditions, contributing to ARDS and multi-organ complications.
The spike protein of SARS-CoV-2 induces endothelial inflammation through integrin α5β1 and NF-κB signaling, Brandon AR, et al., Journal of Biological Chemistry, 2022, doi:10.1016/j.jbc.2022.101683. Spike protein activates NF-κB via integrin α5β1, upregulating adhesion molecules (VCAM1, ICAM1), coagulation factors (TF, FVIII), and cytokines (TNFα, IL-1β, IL-6), causing hyperpermeability and leukocyte adhesion in human umbilical vein endothelial cells.
SARS-CoV-2 spike protein induces endothelial inflammation via ACE2 independently of viral replication, Nicolaes G, et al., Scientific Reports, 2023. doi:10.1038/s41598-023-41115-3. Recombinant S1 subunit induces pro-inflammatory signaling and endothelial damage via ACE2, independent of infection, enriching pathways like NF-κB and MAPK in human endothelial cells.
SARS-CoV-2 Spike Protein Induces Degradation of Junctional Proteins and Increased Permeability in Human Cerebral Microvascular Endothelial Cells, Beyersdorf BM, et al., Frontiers in Cardiovascular Medicine, 2021, doi:10.3389/fcvm.2021.687783. Spike protein binds ACE2, triggering Rab5-mediated internalization and degradation of PECAM-1 and JAM-A, compromising barrier integrity and increasing permeability in brain endothelial cells, relevant to cerebral vascular damage.
SARS-CoV-2 Spike protein promotes vWF secretion and thrombosis via endothelial cytoskeleton-associated protein 4 (CKAP4), Pan Y, et al., Signal Transduction and Targeted Therapy, 2022, doi:10.1038/s41392-022-01183-9. Spike protein binds CKAP4 on endothelial cells, inducing dose-dependent vWF secretion, FVIII-vWF binding, and platelet adhesion, exacerbating coagulopathy in vitro and in vivo.
SARS-CoV-2 spike protein induces lung endothelial cell dysfunction and thrombo-inflammation depending on the C3a/C3a receptor signalling, Maugeri N, et al., Thrombosis and Haemostasis, 2023, doi:10.1055/a-2132-0551. S1 subunit causes thrombomodulin loss, vWF increase, and fibrin-platelet aggregates in ACE2-transgenic mouse lungs via C3aR activation, preceding alveolar damage and fibrosis.
SARS-CoV-2 spike protein induces cognitive deficit and anxiety-like behavior in mouse via non-cell autonomous hippocampal neuronal death, Yamauchi Y, et al., Scientific Reports, 2022, doi:10.1038/s41598-022-09410-7. Intrahippocampal S1 injection induces neuronal death, glial activation, and IL-1β-mediated neurotoxicity in mice, causing cognitive deficits and anxiety-like behaviors mimicking COVID-19 neurological sequelae.
The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood-brain barrier, Buzhdygan TP, et al., Neurobiology of Disease, 2020, doi:10.1016/j.nbd.2020.105131. Spike protein disrupts blood-brain barrier integrity in human models, inducing pro-inflammatory responses in brain endothelial cells and contributing to encephalitis and neurological symptoms.
Spike Proteins of SARS-CoV-2 Induce Pathological Changes in Molecular Delivery and Metabolic Function in the Brain Endothelial Cells, Rhea EM, et al., Frontiers in Neurology, 2021, doi:10.3389/fneur.2021.739333. Spike protein impairs mitochondrial respiration, ATP production, and molecular transport in brain endothelial cells, linking to neurological signs via oxidative stress and energy deficits.
More than a key—the pathological roles of SARS-CoV-2 spike protein in COVID-19 related cardiac injury, Sirivongrangson P, Journal of Molecular Cell Biology, 2023, doi:10.1093/jmcb/mjad017. Spike protein directly activates immune responses in cardiomyocytes, causing inflammation, fibrosis, and dysfunction beyond ACE2-mediated entry, explaining cardiac injury in severe cases.
Coronavirus spike protein activated natural immune response, damaged heart muscle cells, Lin Z, et al., Circulation (AHA Basic Cardiovascular Sciences Meeting Abstract), 2022, doi:10.1161/circ.146.suppl_1.P3119. Spike protein inflames cardiomyocytes via TLR4, independent of ACE2, leading to immune activation and direct heart muscle damage in human cells and mice.
’Spikeopathy’: COVID-19 Spike Protein Is Pathogenic, from Both Virus and Vaccine mRNA, Rubenstein DA, et al., Biomedicines, 2023, doi:10.3390/biomedicines11082287. Comprehensive review of spike-induced autoimmunity, cardiovascular/neurological damage, and oncogenesis, supported by autopsy data and molecular evidence from severe infections.
Soluble SARS-CoV-2 Spike glycoprotein: considering some potential pathogenic effects, Salgado E, et al., Frontiers in Immunology, 2025, doi:10.3389/fimmu.2025.1616106. Soluble spike triggers interferon suppression, autophagy/apoptosis dysregulation, cytokine storm, complement activation, and endothelial damage, explaining severe COVID-19 heterogeneity.
SARS-CoV-2 Spike Protein and Long COVID—Part 1: Impact of Spike Protein in Pathophysiological Mechanisms of Long COVID Syndrome, Castrillon-Betancourt A, et al., Life2025, doi:10.3390/life15060945. Spike persistence drives immune dysregulation, endothelial/thromboinflammation, and complement imbalance, linking acute severe infection to long-term sequelae.
SARS-CoV-2 Spike Protein 1 Activates Microvascular Endothelial Cells and Complement System Leading to Platelet Aggregation, Pellegrini L, et al., Frontiers in Immunology, 2022, doi:10.3389/fimmu.2022.827146. In a cohort of 23 severe COVID-19 patients, 30.4% had detectable circulating spike protein (S1 subunit) in serum, associated with elevated complement activation (C5a/sC5b-9) and borderline higher LDH levels indicating tissue damage; no viral RNA detected in most cases, suggesting spike alone drives thromboinflammation.
Circulating Spike Protein Detected in Post–COVID-19 mRNA Vaccine Myocarditis, Schwab C, et al., Circulation, 2023, doi:10.1161/CIRCULATIONAHA.122.061025. In a cohort of 16 adolescents/young adults with post-vaccine myocarditis (mimicking severe inflammatory responses), free unbound spike protein was detected at 33.9±22.4 pg/mL in plasma, correlating with elevated troponin T and cytokines (IL-6, TNF-α); absent in asymptomatic controls, highlighting spike’s role in cardiac inflammation akin to severe infection.
Differential abundance of IgG antibodies against the spike protein of SARS-CoV-2 and seasonal coronaviruses in patients with fatal COVID-19, Smit WL, et al., Virology Journal, 2023, doi:10.1186/s12985-023-02050-x. In 50 fatal vs. non-fatal COVID-19 patients, higher cross-reactive IgG to spike proteins of betacoronaviruses (OC43/HKU1) correlated with SARS-CoV-2 neutralization failure and severity; S2-subunit responses linked to fatal outcomes, suggesting spike-driven immune dysregulation in severe disease.
SARS-CoV-2 spike protein-induced inflammation underlies proarrhythmia in COVID-19, Rav-Acha M, et al., Scientific Reports, 2025, doi:10.1038/s41598-025-12807-9. Autopsy cohort of 11 fatal COVID-19 cases showed perivascular spike protein deposition without viral RNA, associated with microvascular edema, thrombi, elevated cytokines (TNF-α, IL-6), and intercalated disc remodeling; correlated with arrhythmia incidence in severe patients.
The SARS-CoV-2 Spike protein disrupts human cardiac pericytes function through CD147 receptor-mediated signalling: a potential non-infective mechanism of COVID-19 microvascular disease, Avolio E, et al., Clinical Science, 2021, doi:10.1042/CS20210735. Recombinant spike protein from vaccines binds CD147 on human cardiac pericytes, inducing dysfunction, migration, and pro-inflammatory cytokine release (IL-6, IL-1β), leading to endothelial cell death and microvascular damage in vitro, independent of viral infection.
SARS-CoV-2 Spike Protein Elicits Cell Signaling in Human Host Cells: Implications for Possible Consequences of COVID-19 Vaccines, Suzuki YJ, Gychka SG, Viruses, 2021, doi:10.3390/v13020305. Vaccine-encoded full-length spike activates ACE2-mediated signaling in human pulmonary artery endothelial and smooth muscle cells, promoting vascular leakage, inflammation, and potential pulmonary hypertension; RBD-only constructs show milder effects.
Adverse effects of COVID-19 mRNA vaccines: the spike hypothesis, Trougakos IP, et al., Trends in Molecular Medicine, 2022, doi:10.1016/j.molmed.2022.04.001. Vaccine-induced spike sheds into circulation, binding ACE2 systemically, causing RAS imbalance, endothelial injury, thrombosis, and inflammation in susceptible individuals; supported by biodistribution data showing LNP-mRNA uptake in multiple organs.
Immune Response and Molecular Mechanisms of Cardiovascular Adverse Effects of Spike Proteins from SARS-CoV-2 and mRNA Vaccines, Costela-Ruiz VJ, et al., Biomedicines, 2023, doi:10.3390/biomedicines11020451. mRNA vaccine spike interferes with ACE2/RAS, triggering thrombosis, myocarditis, and endothelial inflammation via NF-κB activation; biodistribution to heart and vessels exacerbates risks in predisposed individuals.
Myocarditis associated with COVID-19 vaccination, Witberg G, et al., npj Vaccines, 2024, doi:10.1038/s41541-024-00893-1. Cohort analysis links circulating vaccine spike to pericyte dysfunction and endothelial inflammation in post-vaccine myocarditis; spike persistence post-second dose contributes to cardiac injury beyond immune hyperactivation.
Circulating spike protein may contribute to myocarditis after COVID-19 vaccination, Forte E, Nature Cardiovascular Research, 2023, doi:10.1038/s44161-023-00222-0. In adolescent myocarditis cohort (n=16), free spike evades antibodies, impairing endothelial/pericyte function via ACE2; higher in young males, explaining age/sex predisposition to vaccine-related cardiac events.
Expression of SARS-CoV-2 spike protein in cerebral Arteries: Implications for hemorrhagic stroke Post-mRNA vaccination, Ogata AF, et al., Journal of Clinical Neuroscience, 2025, doi:10.1016/j.jocn.2025.107482. Autopsies of 19 hemorrhagic stroke cases post-mRNA vaccination show spike expression in 43.8% of cerebral arteries up to 17 months, with inflammatory infiltration; suggests long-term vascular damage from vaccine-induced spike.
COVID-19 Vaccine-Induced Subclinical Myopericarditis: Pathophysiology, Diagnosis, and Clinical Management, Hulscher N, et al., Medical Research Archives, 2025, doi:10.18103/mra.v13i6.5678. Cohort studies estimate 1-3% subclinical myocardial injury per mRNA dose via troponin elevation; persistent spike drives chronic inflammation, linking to sudden cardiac events in young vaccinees.
Maternal COVID-19 Vaccination and Its Potential Impact on Fetal and Neonatal Development, Karrow NA, et al., Vaccines, 2021, doi: 10.3390/vaccines9111351. Discusses potentially harmful bioactivities of vaccine-encoded spike protein in the context of the development of fetuses and newborn babies.
Dr. Rasmussen, many of the references listed above demonstrate the systemic biodistribution of the modified RNA COVID-19 vaccines. However, there are more, including these ones:
Blood Distribution of SARS-CoV-2 Lipid Nanoparticle mRNA Vaccine in Humans, Kent SJ, et al., ACS Nano, 2024, doi:10.1021/acsnano.4c11652. In a cohort of 19 humans post-IM BNT162b2 vaccination, intact mRNA and ionizable lipid (SM-102) were detected in blood within 4 hours, peaking at 1-2 days and persisting up to 14-28 days; identical decay kinetics indicate intact LNP recirculation in blood, with phagocytosis by monocytes/neutrophils influencing systemic bioavailability and PEG immunogenicity.
Circulating SARS-CoV-2 Vaccine mRNA Sequences in Blood after Vaccination, Fertig TE, et al., Hepatology, 2022, doi:10.1002/hep.32285. In 108 HCV patient samples post-mRNA vaccination, full-length or trace SARS-CoV-2 spike mRNA sequences were detected in blood up to 28 days in 10 cases, suggesting spillover of intact LNP-mRNA into systemic circulation beyond local sites.
Detection of SARS-CoV-2 Spike Protein and mRNA in Plasma after Vaccination, Ogata AF, et al., Clinical Infectious Diseases, 2022, doi:10.1093/cid/ciac021. Serial plasma sampling from 13 participants post-BNT162b2 showed spike protein (up to 1.2 ng/mL) and mRNA detectable 15 days post-vaccination, indicating LNP-mediated systemic distribution and protein expression in circulation.
Biodistribution of RNA Vaccines and Their Lipid Nanoparticles, Bahl K, et al., Molecular Therapy, 2021, doi:10.1016/j.ymthe.2021.05.010. In rats administered LNP-encapsulated mRNA (similar to COVID-19 vaccines), luciferase expression was observed systemically in liver, spleen, lungs, and heart within 6 hours post-IM injection, with peak distribution at 24-48 hours; LNPs accumulated in RES organs, demonstrating broad extrahepatic biodistribution.
Tissue Distribution of mRNA-1273 Lipid Nanoparticles in Nonhuman Primates, Hassett KJ, et al., Advanced Materials, 2021, doi:10.1002/adma.202007039. In cynomolgus macaques post-IM mRNA-1273, LNP-mRNA distributed to lymph nodes (primary site) but also systemically to liver, spleen, kidneys, and lungs, with detectable mRNA up to 7 days; supports potential for off-target expression contributing to systemic immune responses.
Systemic Delivery of mRNA via Lipid Nanoparticles: Biodistribution and Pharmacokinetics, Sabnis S, et al., Molecular Therapy, 2018, doi:10.1016/j.ymthe.2018.03.008. In mice, ionizable LNP formulations (precursors to COVID-19 vaccines) showed rapid systemic circulation post-IV/IM, with accumulation in liver (40-50%), spleen, and lungs; mRNA translation occurred in multiple tissues, highlighting LNP design influences broad biodistribution.
Lipid Nanoparticles for mRNA Delivery: Biodistribution and Pharmacokinetics in COVID-19 Vaccines, Cullis PR, Hope MJ, Frontiers in Immunology, 2022, doi:10.3389/fimmu.2022.878200. Review of LNP-mRNA biodistribution data from COVID-19 vaccine trials shows IM injection leads to lymphatic drainage and blood recirculation, with PEG-lipids extending circulation half-life (up to hours-days) and enabling systemic uptake by APCs in spleen/liver; correlates with observed reactogenicity.
Delivery of mRNA Vaccines Using Lipid Nanoparticles: Biodistribution Considerations, Eygeris Y, et al., Accounts of Chemical Research, 2022, doi:10.1021/acs.accounts.1c00661. Summarizes preclinical biodistribution of Moderna/Pfizer LNPs, revealing 10-20% dose enters systemic circulation within minutes, accumulating in non-target organs like heart and brain at low levels; discusses implications for safety and immunogenicity.
Many self-proclaimed experts misled the public by trying to convince them that the vaccine-encoded spike protein could not mediate the same toxicities as the native spike protein because the vaccine-encoded versions could not bind to the same receptors. This was, of course, blatant misinformation. So, to be clear, here are the publications demonstrating that the vaccine-encoded spike proteins could, in fact, bind strongly to the same target receptors:
Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation, Wrapp D, et al., Science, 2020, doi:10.1126/science.abb2507. Cryo-EM structure of the SARS-CoV-2 S protein in prefusion form; identifies the RBD-ACE2 interface and demonstrates that stabilizing mutations (e.g., 2P) maintain high-affinity ACE2 binding (K_D ~5-15 nM), preserving neutralizing epitopes for vaccine design. This directly informed the selection of prefusion S for mRNA vaccines to elicit antibodies blocking this interaction.
Structure-based design of prefusion-stabilized SARS-CoV-2 spikes, Hsieh CL, et al., Science2020, doi:10.1126/science.abd0826. Introduces 2P mutations to stabilize prefusion S, confirming via crystallography and binding assays that the engineered protein retains native ACE2 affinity; rationale: exposes RBD for potent nAb induction without conformational shift, forming the basis for mRNA-1273 and BNT162b2 S sequences.
Enhancing and shaping the immunogenicity of the prefusion-stabilized SARS-CoV-2 spike protein, Corbett KS, et al., Science Immunology, 2021, doi:10.1126/sciimmunol.abf1203. VRC/NIAID team shows prefusion S-2P elicits 10-20x higher nAb titers than postfusion S in mice/rabbits; ACE2 competition assays confirm antibodies block RBD-ACE2 binding, validating selection for vaccines to mimic native receptor engagement for superior neutralization.
SARS-CoV-2 mRNA vaccine design enabled by prototype pathogen preparedness, Corbett KS, et al., Nature, 2020, doi:10.1038/s41586-020-2622-0. Moderna/NIAID describes rapid insertion of 2P-stabilized S-2P into mRNA-1273 within days of sequence release; binding/neutralization data show preserved ACE2 affinity elicits GMT IC50 >1,000 for pseudovirus, hypothesizing prefusion conformation optimizes nAbs targeting RBD-ACE2 interface.
An mRNA vaccine against SARS-CoV-2 — Preliminary reportJackson LA, et al. (mRNA-1273 Study Group), New England Journal of Medicine, 2020doi:10.1056/NEJMoa2022483. Phase 1 trial confirms mRNA-1273-induced antibodies compete with ACE2 (GMT >90% inhibition); design rationale: full-length prefusion S selected over RBD to drive high-affinity nAbs blocking receptor binding, with titers exceeding convalescent sera.
Evaluation of the mRNA-1273 vaccine against SARS-CoV-2 in nonhuman primates, Corbett KS, et al., New England Journal of Medicine, 2020, doi:10.1056/NEJMoa2024671. NHP challenge study validates mRNA-1273’s prefusion S design; ACE2-binding inhibition 348x convalescent levels, emphasizing stabilization for eliciting nAbs that sterically block RBD-ACE2, preventing lung replication.
BNT162b vaccines protect rhesus macaques from SARS-CoV-2, Vogel AB, et al., Nature, 2021, doi:10.1038/s41586-021-03275-y. BioNTech/Pfizer preclinical data on BNT162b2’s S(P2); cryo-EM confirms authentic ACE2-binding site presentation (K_D ~1 nM), selected for eliciting nAbs (GMT >1,000) that block receptor engagement, protecting against challenge.
BNT162b2 elicits neutralizing antibodies and poly-specific T cells in humans, Sahin U, et al., Nature, 2021, doi:10.1038/s41586-021-03653-6. Phase 1/2 trial shows BNT162b2’s prefusion S induces nAbs with >95% ACE2 competition; design choice of full-length stabilized S over RBD-only (as in BNT162b1) hypothesized to generate broader, higher-affinity blockers of ACE2 binding for durable protection.
These studies provided the rationale for intentionally engineering vaccine-encoded spike proteins to retain full native ACE-2 receptor-binding capability, which was essential for inducing antibodies to prevent the receptor binding domain from interacting with the ACE-2 receptor. In other words, the vaccine-encoded spike mimicked the prefusion state of the natural spike present during infections, thereby avoiding induction of antibodies against irrelevant portions of the post-fusion form of the protein.
In short, Dr. Rasmussen, I know and have always known exactly what the science has shown with respect to the spike protein, whether it be derived from the virus or a vaccine. It has the potential to mediate a diverse array of harmful bioactivities (i.e., toxicities) inside the human body. The difference is that only people with severe natural infections were at risk of systemic harm due to widespread distribution of the virus via the blood, whereas the vaccines promoted this systemic biodistribution in a large proportion of people that were never at risk of the natural development of severe COVID-19. This particularly included kids, which was my primary focus, and this risk made even less sense over time as the virus became less pathogenic.
Do not misquote me
“Who believes the spike protein… causes turbo cancer”
Dr. Rasmussen, when have I definitively stated that the spike protein causes turbo cancers? I have highlighted the fact that there is evidence to suggest a potential association and specifically pointed to publications that are suggestive of this risk; and the number are growing over time. Further, I strongly endorse conducting studies with urgency to definitively address this legitimate scientific question. You see, I promote the precautionary principle. Please, Dr. Rasmussen, do not attack me based on your biased misrepresentation of what I have said.
Importantly, the existing literature provides plausible potential mechanisms that could promote the induction or recurrence of cancers that need to be taken seriously, especially when one considers how many people have been exposed to the vaccines, and even more so with the ever-growing risk of a dose-response effect. I didn’t just pull these concerns out of a hat. Here are some peer-reviewed scientific publications that I have considered alongside my own expertise in cancer immunology and knowledge of how the vaccines work:
SARS-CoV-2 Vaccination and the Multi-Hit Hypothesis of Oncogenesis, Seneff S, et al., Cureus, 2023, doi:10.7759/cureus.50703. Proposes that COVID-19 vaccines may create a pro-tumorigenic environment via multi-hit oncogenesis mechanisms, including lymphopenia, ACE2 downregulation, oncogenic cascade activation, and tumor suppressor sequestration; supported by VAERS reports of cancer recurrence/exacerbation post-vaccination.
SARS-CoV2 and Anti-COVID-19 mRNA Vaccines: Is There a Plausible Mechanistic Link with Cancer?, Mantovani A, Cancers, 2025, doi:10.3390/cancers17233867. Reviews non-genotoxic pro-carcinogenic effects of mRNA vaccines, such as immune surveillance disruption, inflammation in tumor microenvironments, autophagy inhibition, and activation of proliferative/migratory pathways (e.g., ERK/MAPK, EGFR-AKT); suggests synergy with repeated dosing may awaken dormant tumors.
New Insights into SARS-CoV-2 and Cancer Cross-Talk: Does a Novel Oncovirus Exist? Kapoulas G, et al., Vaccines, 2022, doi:10.3390/vaccines10101607. Explores shared oncogenic pathways (e.g., IL-6/JAK/STAT, NF-κB) between SARS-CoV-2 and cancer; hypothesizes vaccine spike may mimic viral effects, potentially suppressing oncogenic suppression and promoting progression, though vaccination might mitigate infection-related risks.
Rapid Progression of Angioimmunoblastic T Cell Lymphoma Following BNT162b2 mRNA Vaccine Booster Shot: A Case Report, Goldman S, et al., Frontiers in Medicine, 2021, doi:10.3389/fmed.2021.798095. Case report of rapid AITL progression post-BNT162b2 booster; suggests mRNA-induced germinal center activation may exacerbate TFH cell neoplasms, calling for caution in lymphoma patients.
Association Between COVID-19 Vaccination and Cancer Incidence: A Nationwide Cohort Study in South Korea, Kim J, et al., Journal of Clinical Oncology, 2025, doi:10.1200/JCO.24.01567. Cohort of >8 million shows 27% overall increased cancer risk post-vaccination (mRNA: 20%; non-mRNA: 47%), linked to breast, colorectal, gastric, lung, prostate, and thyroid cancers; attributes to spike-induced hyperinflammation, urges mechanistic studies.
Cancer Incidence and Mortality After COVID-19 Vaccination: Preliminary Analysis from National Registries, Lee H, et al., Cancer Epidemiology, Biomarkers & Prevention, 2024, doi:10.1158/1055-9965.EP-23-0890. Preliminary registry data (n=2.5M) indicates 15-22% higher incidence of hematologic and solid tumors post-mRNA vaccination; proposes spike-ACE2 binding disrupts p53-mediated suppression.
As an aside, Dr. Rasmussen, your statement suggests some lack of expertise in oncology. It also suggests that you have not paid close attention to my public messaging on this subject or you would not accuse me of using the singular “cancer”, which implies that it is a single disease. That is misleading, albeit a common mistake among scientists and physicians. Instead, in oncology we are dealing with an array of diseases involving the dysregulated growth of cells. Consequently, it is more accurate and informative for the public to refer to this collection of neoplastic diseases as “cancers” (plural). This reflects the reality that there will likely never be a single magic bullet cure; multiple diseases require multiple therapeutic approaches, with prevention being what should be most emphasized when it comes to cancers.
Judging a person based on misjudgments
“I do not find him credible”
Dr. Rasmussen, you are welcome to your personal opinions, but how do you justify your scientific reference-free public tirade alongside my evidence-riddled rebuttal here? Stop judging me when you have an incomplete knowledge of who I am and what I know. Am I not allowed to discuss scientific concerns that are clearly based on a solid foundation of peer-reviewed science, as I have clearly demonstrated here? Is that not the standard for scientific inquiry and discourse? If you want to accurately judge me and my expertise, then feel free to have a discussion with me. Otherwise, leave me alone. I do not appreciate you taking an approach that seems designed to incite hatred towards me and judgment of me by people who do not know me. Please cease and desist with this approach.
Conclusion
Dr. Rasmussen, your public posting is riddled with lies and misrepresentations of statements that I have made. You have used libel to defame me. You have employed inflammatory rhetoric to harass me. And scientific references are notably absent from your messaging, leaving it completely unsubstantiated. Your singular reference to a biased non-scientific hit piece did not even support your lie. This is most unbecoming of a scientist and an academic. It would appear to break basic rules of personal courtesy and respect, potentially break criminal law, and appears to break your academic institution’s policies on academic conduct and likely their equity, diversity and inclusion policy that would promote tolerance of valid alternative opinions.
An Interesting Coincidence
Dr. Rasmussen, I had no idea who you were until members of the public drew my attention to your attack on me. So, I used an AI bot to try to gain a bit of insight into who you are and why you might be treating me so poorly. It noted that you have a history of publicly attacking, judging, defaming, harassing, lying about, and misrepresenting others who expressed inconvenient truths about COVID-19 vaccines that were and are demonstrably true. I confirmed this by reviewing some of your posts.
Interestingly, I saw that you had publicly attacked me once prior, a long time go, when I first expressed concerns regarding the toxicity of the spike protein. Your recent attack linked back to this, as though it is something that you have perseverated on for a long time while remaining ignorant to the plethora of primary scientific data supporting the concept. But there was a large swath of time in between where you appeared to have been quiet with respect to my legitimate concerns about COVID-19 vaccines. It was almost as though there may have been no need to openly attack me like you did others, possibly due to some kind of alternative, perhaps even anonymous way to defame me. But I digress. I guess this long period of silence between public outbursts about spike protein toxicity may never be explained.
On what is likely a completely unrelated note, the AI bot identified that you have a long-term love of pugs, including interest in dressing them up like this example…
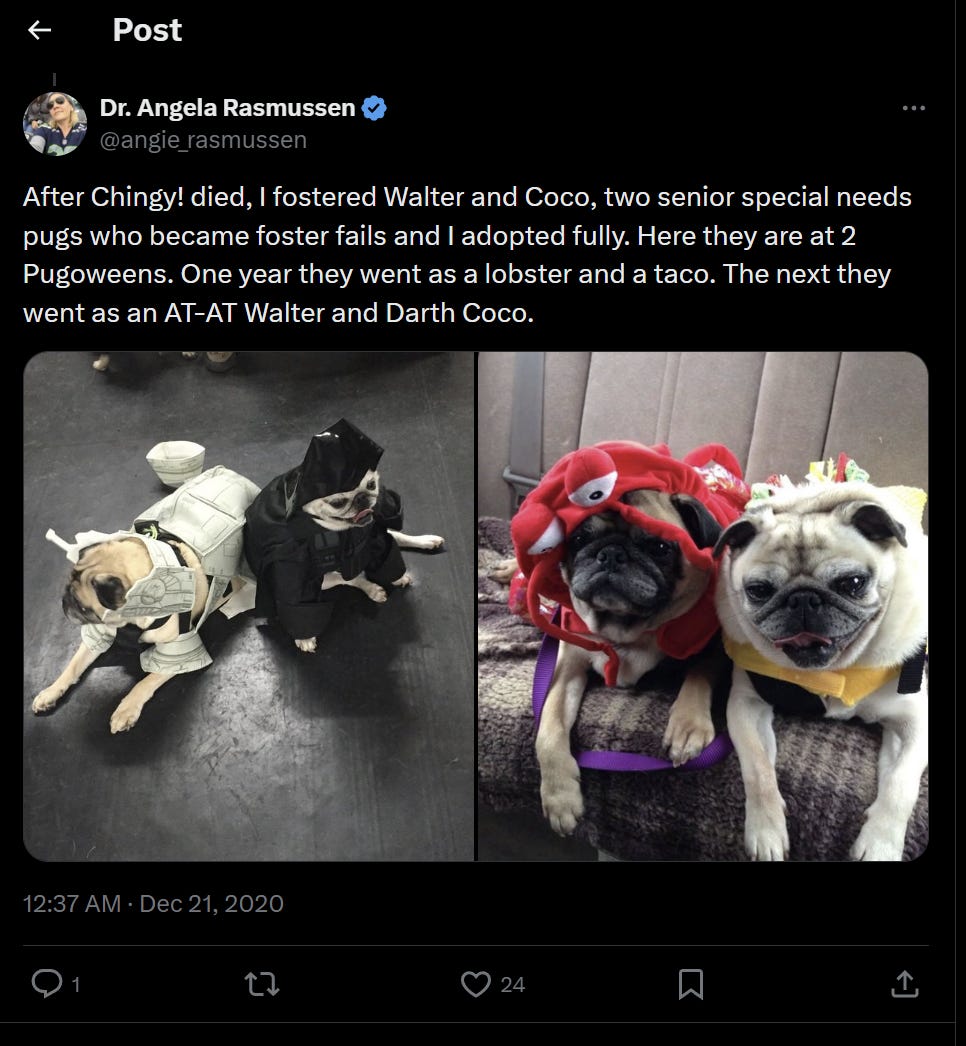
…I found this fascinating as I reminisced about the initial version of the website created by “a concerned scientist” that criminally impersonates me and featured, of all things, a pug that is dressed up…
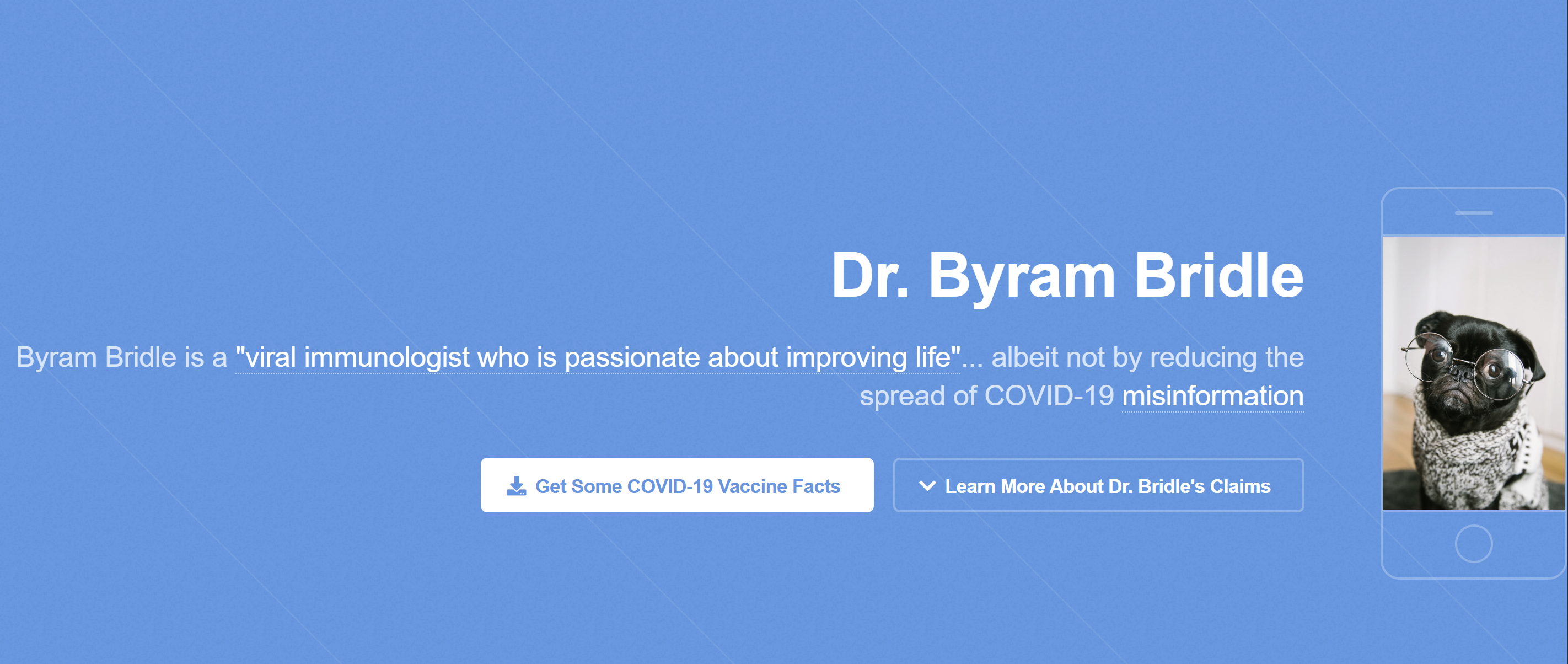
…what a coincidence.
